
In this session, we will discuss the clinical localisation of sensory symptoms. We will learn the sensory pathways and study the clinical features of lesions involving these pathways. At the of this session, we will be able to tell
1. The paths of various sensory modalities and the arrangement of fibres in each.
2. Clinical features of lesions at different sensory levels and
3. To localise from the patient’s symptomatology wherein the sensory pathway is the lesion.
The primary modalities of sensation, including touch, pain, temperature, JPS, and vibration, are recognised at the level of the opposite thalamus. So all these sensations are carried to the opposite thalamus by the sensory pathways. The fine touch, vibration and JPS ascend in the posterior column- medial lemniscus pathway, while the pain and temperature through the lateral spinothalamic tract. From the thalamus, they are then projected to the sensory cortex in the postcentral gyrus through the posterior limb of the internal capsule. The sensory cortex helps identify the cortical sensations, namely tactile localisation, two-point discrimination, graphesthesia, stereognosis and sensory attention.
The sensory symptoms can be classified into two, positive sensory symptoms and negative sensory symptoms. The positive sensory symptoms include paresthesia, radicular pain, allodynia, girdle like sensation etc. Paresthesia is an abnormal spontaneous sensation felt in the absence of a stimulus. Allodynia is a pain response to a stimulus that is not usually painful. The negative sensory symptom is one where a normal sensation is not perceived. Negative sensory symptoms include anaesthesia, where all modalities of sensation are impaired and the washbasin sign, which is the tendency to fall on closing eyes, the symptom equivalent to the positive Romberg sign.
Sherrington classified the sensory system into exteroceptive, interoceptive and proprioceptive. The exteroceptive sensations provide information about the external environment. It includes general somatosensory functions like touch, pain and temperature, and special senses, namely vision, hearing, taste, and vestibular. The Interoceptive sensation provides information about the visceral functions, including blood pressure, blood composition, etc. Proprioception provides information about the orientation of the body and limbs in space.
The exteroceptive sensation
The exteroceptive sensation originates in the peripheral receptors in response to external stimuli. There are four main general somatic sensations. They are pain, temperature, touch and joint position sense.
Touch
The receptors for touch sensation include free nerve endings, Merkel cell endings, and encapsulated nerve endings such as Meissner’s corpuscles, Pacinian corpuscles and Ruffini endings. The Pacinian corpuscles act as rapidly adapting mechanoreceptors and are especially sensitive to high-frequency vibration. Others are slowly adapting mechanoreceptors. Light touch sensation is conveyed over large myelinated peripheral nerve fibres to unipolar DRG cells. Tactile sensation follows several different pathways within the central nervous system.
The central processes of DRG enter the spinal cord via the medial division of the posterior roots. It then projects through the ipsilateral posterior column to reach the cervicomedullary junction. The fibres from the sacral area will be arranged most medially in the posterior column. The lumbar will be just lateral, followed by the thoracic. The cervical fibres are lateral most as they are the last to enter while going up. The second-order neuron lies in the cuneatus and gracillis nucleus at the cervicomedullary junction. All the fibres below T8 are grouped in the fasciculus gracilis; fibres above T8 form the fasciculus cuneatus. The second-order neurons in the nucleus cuneatus and gracillis project to the opposite thalamus. They immediately sweep anteriorly as the internal arcuate fibres and cross over to the opposite side at the cervicomedullay junction. It ascends in the brainstem as the medial lemniscus to reach the thalamus. The Third-order neuron lies in the ventral posterolateral nucleus of the thalamus and project to the sensory cortex.
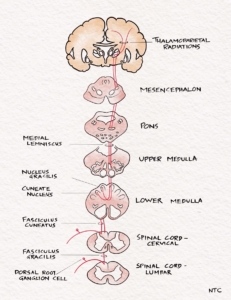
Fig -1 The fine touch pathway
The fibres that carry crude touch synapse after entering the posterior horn. The axons of the second-order neuron cross to the opposite anterior spinothalamic tract and reaches the thalamus.
Few other tactile fibres have a synapse in the posterior horn. Then they ascend in the dorsolateral funiculus to the lateral cervical nucleus at C1-C2. The fibres then decussate and join the medial lemniscus. Thus tactile sensation reaches the opposite thalamus through different routes. This accounts for the poor localising value of touch in spinal cord disorders and is lost only in extensive axial lesions.
The touch sensation from the face will reach the principal sensory nucleus of the trigeminal nerve. It crosses over to the opposite side and ascends along the medial lemniscus as the ventral trigeminal tract. It ends in the ventral posteromedial nucleus of the thalamus. The ventral trigeminal tract also contains fibres from the opposite spinal nucleus of the trigeminal nerve that carries the pain sensation of the face. Some fibres will not cross to the opposite side and form the dorsal trigeminal tract. The ventral and dorsal trigeminal tract is called the trigeminothalamic tract, trigeminal lemniscus, or Quinto thalamic tract.
Pain and temperature
The superficial pain sensation arises in nociceptors—free nerve endings in the skin and mucous membranes. Thermoreceptors for heat and cold sensations are free nerve endings in the dermis. Pain and thermal sensation are carried along small myelinated A-delta and unmyelinated C nerve fibres to the dorsal root ganglion. First-order neurons in DRG psudounipolar cells enter through the lateral part of the dorsal root to reach the Lissauer’s tract, where they ascend for one or two segments. The axons leave Lissauer’s tract, enter the posterior grey horn, and synapse in the laminae I, II, and V. The second-order neuron crosses to the opposite side through the anterior commissure. It forms the lateral spinothalamic tract, which ascends through the spinal cord and brainstem to reach the opposite thalamus. In the past, anatomists thought the anterior spinothalamic tract carried crude touch, and the lateral spinothalamic tract carried pain and temperature. Current evidence suggests all these modalities are carried in both tracts together, called the anterolateral system.
The fibres from the sacral area are arranged most laterally in the lateral spinothalamic tract, followed by lumbar, thoracic and cervical. The cervical fibres, which are last to arrive while going up, are the medial most. So an intramedullary lesion like syringomyelia will affect laterally placed sacral fibres last. The third-order neuron in the thalamus projects to the sensory cortex in the postcentral gyrus through the posterior limb of the internal capsule.
The pain sensation from the face is carried by the spinal tract of the trigeminal nerve and synapses in the adjacent nucleus of the spinal tract. The second-order neuron crosses over to the opposite side, lie adjacent to the medial lemniscus and lateral spinothalamic tract and forms the trigeminothalamic tract, which terminates in the VPM nucleus of the thalamus from where it is projected to the sensory cortex.
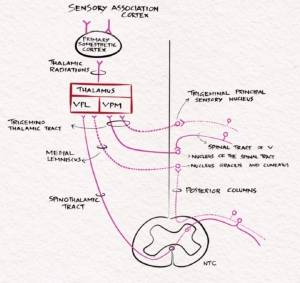
Fig-2 Schematic diagram of the touch and pain pathway
The proprioceptive sensation
The proprioceptive sensation arises from the body’s deeper structures: bones, ligaments, muscles, and joints. Proprioception refers to either the sense of the position of a body part or the motion of a body part. Proprioception has both a conscious and an unconscious component. The conscious element is carried by the posterior column and the unconscious by the spinocerebellar tract.
The primary receptor for proprioception or kinesthesia is the muscle spindle. The pathway of the conscious proprioception is the same as that of light touch through the posterior column-medial lemniscus. The proprioceptive impulses from the head and neck travel through cranial nerves and end in the mesencephalic nucleus of the trigeminal nerve. From there, they reach the opposite thalamus through the medial lemniscus.
Vibration sense or pallesthesia
The receptors of vibration are rapidly adapting mechanoreceptors like Pacinian corpuscle. The Merkel disk receptors and Meissner corpuscles respond best to relatively low frequencies while Pacinian corpuscles to higher frequencies. Impulses are relayed with the proprioceptive and touch sensations through large, myelinated nerve fibres. They enter the spinal cord through the medial division of the posterior root. It ascends in the spinal cord through multiple pathways, namely posterior column-medial lemniscus and dorsolateral funiculus.
The fibres in the dorsolateral funiculus may be the most crucial pathway subserving vibratory sensation in man. Axons of the second-order neurons in the posterior horn ascend in the spinocervical tract in the ipsilateral dorsolateral funiculus. It terminates in the lateral cervical nucleus. Axons from neurons in the lateral cervical nucleus cross in the anterior commissure and ascend to the medulla, where they join the ML. Though some JPS fibres also ascend through this pathway, this divergence of the position sense and vibration sense pathways partially explains the dissociation occasionally encountered clinically between JPS and vibration. For example, vibration loss may be worse than position sense loss in subacute combined degeneration, conversely for tabes dorsalis. Position sense is often impaired with a parietal lobe lesion and vibration preserved
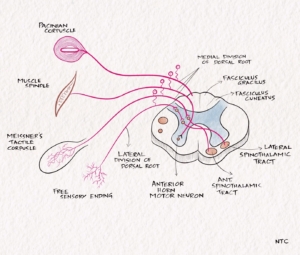
Fig -3 The various sensory receptors and the tracts
Interoceptive sensations
Interoceptive sensations arise from the internal organs. General visceral afferent fibres are found in cranial nerves VII, IX, X, and thoracolumbar and sacral autonomic nerves. Visceral afferent fibres run with autonomic efferent fibres to the viscera. Cell bodies are in the dorsal root and associated cranial ganglia. They convey visceral sensations such as hunger, nausea, sexual excitement, vesical distention, and visceral pain. They ascend in the spinal cord through different routes. Some ascend in the opposite spinothalamic tract. Some travel in the ipsilateral spinothalamic tract. Some ascend for a great distance in the Lissauer tract before synapsing with second-order neurons. Others ascend by long intersegmental fibres in the white matter at the border of the dorsal horn, reaching the hypothalamus and thalamus without decussating. Because of the multiple pathways, the localisation of visceral pain is not usually exact. The visceral afferent sensations may end in the gyms rectus, rather than in the parietal cortex.
Localisation of sensory symptoms
Sensory cortex
As already described, the primary modalities of sensation are recognised at the level of the thalamus. So touch, vibration, JPS, pain and temperature will be intact in a parietal cortex lesion. The sensory fibres are then projected to the sensory cortex in the postcentral gyrus through the posterior limb of the internal capsule. The parietal lobe primarily does cortical sensory processing. The parietal lobe integrates more than one primary sensory modality to recognise a stimulus. Cortical sensations like tactile localisation, two-point discrimination, graphesthesia, and stereognosis will be impaired in the parietal lesion. The patient can have sensory inattention, especially with a right-sided lesion. In sensory inattention, the patient can identify sensation when either side is stimulated separately. On stimulating both sides together, only the right side is appreciated. Rarely lesions of the sensory cortex and sensory projections in the posterior limb of the internal capsule can cause a loss of appreciation of primary modalities of the opposite half of the body as in a thalamic lesion. In that case, the sensory loss may be more in the upper limb than the lower limb, and distal more than proximal as the area of representation in the sensory cortex is more for these regions. Even more rarely, small lesions in the sensory cortex can produce a restricted sensory loss mimicking a nerve or root lesion.
Thalamus
Thalamic lesions produce impairment of all modalities of primary sensation in the opposite half of the body, including the face. A small lesion restricted to the ventral-posterolateral nucleus can cause hemi paresthesia without demonstrable sensory loss. Thalamic lesions can cause sensory perversions like paresthesias, hyperesthesias and allodynia. A patient with a thalamic lesion can later develop a central pain syndrome of the opposite side, a condition called Dejerine Roussey syndrome. Impairment in sensation with intractable pain in the same area is called anaesthesia dolorosa. A parietal opercular lesion can produce a central pain syndrome mimicking a thalamic lesion called pseudothalamic syndrome.
Brainstem
The touch and pain pathways are well separated in the brainstem and spinal cord. So lesions here can affect one tract, sparing the other resulting in dissociated anaesthesia. For example, a medial medullary syndrome produces loss of fine touch, vibration and JPS of the opposite side due to medial lemniscal involvement with sparing of pain and temperature. Another feature of a brainstem sensory syndrome is the involvement of face sensation on the side of the lesion and sensation of the opposite side of the body due to crossed over sensory tract involvement. The classic example is the lateral medullary syndrome, where pain and temperature over the face are lost on the same side due to the long tract of trigeminal involvement. The pain over the opposite side of the body is lost due to lateral spinothalamic tract involvement. Rarely a lower brainstem lesion can produce a sensory level on the trunk.
Spinal cord
In the spinal cord, the touch, vibration and JPS ascend in the ipsilateral posterior column while pain and temperature cross over and ascend on the opposite side lateral spinothalamic tract. Several sensory syndromes can occur with spinal cord lesions, namely, Brown-Sequard, complete transection, central cord syndrome, anterior spinal artery syndrome, conus, conuscauda etc. All of them are discussed in detail in the episodes on the anatomy and localisation of spinal cord disorders. Please hear episode 5 to revise that.
Lesions of the root, plexus and peripheral nerves
Lesions of the root, plexus and peripheral nerves will result in the involvement of all modalities of sensation in the distribution of the particular root or nerve. It is essential to know the dermatomes and sensory supply of peripheral nerves to localise these lesions correctly. These images are given on the episode web page provided in the show notes. The area of actual sensory loss will be much less than that given in the pictures because of the sensory overlap. The patient can have either positive sensory symptoms like paresthesia or hyperesthesia or negative sensory symptom like hypesthesia in the area of involvement.
Polyneuropathies
Multiple peripheral nerves are affected together in polyneuropathies. The classic example is diabetic peripheral neuropathy, with a length-dependent stocking and glove type of peripheral neuropathy. The sensory impairment starts in the sole of the feet symmetrically. It then ascends to the knee and then appears in the arm in a length-dependent manner. Vibration is the first to get involved, often followed later by the other modalities. Some polyneuropathies have a predilection for large myelinated fibres that carry touch, JPS and vibration. Others for small fibres that carry pain and temperature. Sjogren’s syndrome, pyridoxine toxicity, Cisplatin and B12 deficiency affect large fibres more. Hansen’s disease, Tangier’s disease and amyloidosis affect small fibres more. Large fibre neuropathies are associated with reflex loss and, when severe, with motor involvement. Small fibre neuropathies produce burning paresthesia with no motor loss and preserved reflexes. Nerve conduction study may be normal in pure small fibre neuropathy.
Dorsal root ganglionopathy
Autoimmune disorders can affect DRG. The large myelinated fibres are affected more than small fibres. The patient will have paresthesia, generalised areflexia and, if severe, sensory ataxia and pseudoathetosis. It may be associated with malignancy like small cell carcinoma lung and lymphoma, Sjogren syndrome and pyridoxine toxicity.
It’s been more than nine months since we started our podcast. The podcast has been downloaded more than 11000 times from over 93 countries. It is now one of the top 10 international neurology podcasts, according to Feedspot. The link is given in the show notes. It brings great joy to see all the hard work paying off and see that people find it useful. Please continue your valuable support and share it with your friends and colleagues. Kindly write a review and provide a rating on the Apple podcast and Spotify. It will be a big help and will be much appreciated.